
|
| 首页 > 新闻 > 科研新闻 > 正文 | 字号选择: |
| 稻米品质形成的生理基础研究进展 |
|
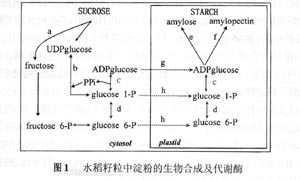
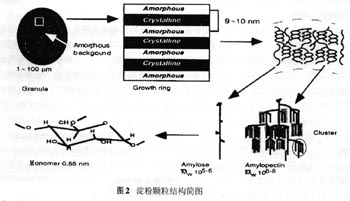
胚乳是水稻籽粒的主要组成部分,占糙米重量的90%,其发育和充实状况不但决定了籽粒的重量,同时也决定了稻米的品质。黄发松等(1998年)认为水稻品质形成过程可以描述为:在遗传特性和环境的作用下,通过籽粒灌浆动态变化来决定其品质的表现,而在水稻籽粒形成过程中酶活性的变化和灌浆速率也有其固有的表现,并认为环境条件对稻米品质的影响是通过影响谷粒胚乳细胞发育及内部生理生化过程等发挥作用的。本文主要从米质形成的生理基础的角度,对国内外有关稻米品质的研究进展进行综述。 1 稻米品质形成期的"源库”特征与垩白的形成 已有的研究表明:评价稻米外观品质的重要指标之一的垩白主要受稻米品质形成期“源库”水平的影响(高如嵩和张嵩午,1994年;钟旭华等,1996年)。垩白形成的一种观点认为,在不利的成熟条件下,源的不足是垩白形成的主要原因(赵式英,1982年)。田代亨和江幡守卫(1975年)研究证明:在抽穗后0~20天内遮光,减少前期的灌浆物质量,垩白米降低;而在抽穗后21~40天内遮光,减少后期的灌浆物质量,垩白米率增加。长户一雄(1958年)研究表明,在抽穗后10天内剪去部分枝梗,增加前期单位颖花的灌浆物质量,垩白米率增加;而在抽穗后15天以后剪去部分枝梗,增加后期单位颖花的灌浆物质量,垩白米率降低。孙义伟和刘宜柏(1990年)研究指出,垩白的形成与籽粒灌浆物质(源)动态有密切关系,平缓线性型灌浆品种其垩白往往较小,而大起大落型垩白较大。邵冬生和唐建军(1987年)研究水稻籽粒灌浆特性与米质的关系时发现,一般情况下,谷粒灌浆前和中期的灌浆速率与总精米率、整精米率、垩白大小、碱解度和胶稠度均呈反相关趋势,而与透明度呈正相关趋势;而灌浆后期的情况则与前和中期表现相反的趋势。大量的育种实践表明,籽粒大小与垩白出现有关,籽粒越大,垩白出现概率越高。贾志宽等(1991年)测得垩白米的容重低于非垩白米,张佩莲等(1995年)研究认为垩白米率与千粒重呈负相关。由此可知,水稻籽粒灌浆的前和后期灌浆物质的水平以及源的动态在一定程度上与稻米的垩白的形成有关,从而影响了稻米的外观品质。 库的活性是指库接受光合产物的能力,增加库活性有利于减少垩白。梁建生等(1993年)研究籽粒发育过程中籽粒库强与其淀粉积累之间的关系发现,强和弱势粒淀粉积累与其胚乳细胞数目密切相关,两者表现出明显的同步性。蔡一霞等(2002年)发现,不同土壤水分处理稻株的颖花绿叶量(绿叶面积与籽粒总数之比,单位为:cm2/颖花)、叶绿素含量和籽粒含水量等生理指标有明显变化,低限土壤水势≤-30 kPa下,灌浆中后期(花后25~35天)的颖花绿叶量和叶绿素含量的相对差值表现出明显的负效应,籽粒失水较快,从而影响米质形成期光合生产和籽粒中淀粉的积累,最终导致米质变劣,整精米率显著下降,垩白粒率和垩白度显著增加。 2 稻米品质形成过程中的关键酶 稻米淀粉是精米的主要成分,稻米品质的形成实质上可以看作是籽粒中淀粉的生物合成和累积的过程,而这一过程发生在淀粉体中,并在一系列酶的催化作用下形成(图1)。根据已有的研究来看,籽粒淀粉形成过程中,蔗糖合成酶、ADP-葡萄糖焦磷酸化酶、Q酶、可溶性淀粉合成酶及淀粉粒形成酶等与稻米品质的关系最密切,其中Q酶是水稻胚乳淀粉生物合成的关键酶,它可催化α-1,6葡萄糖苷键的合成,在直链上产生分支而形成支链淀粉(Nakanuma等,1992年)。近年来的研究发现淀粉去分支酶在淀粉颗粒形成中有着重要的作用,正常的支链淀粉的分支模式是由淀粉分支酶(starch branching enzyme,SBE)和去分支酶(debranching enzyme,DBE)平衡作用的结果,植物糖原是在DBE缺失或活性降低的情况下,只通过SBE催化作用而形成的(Nakanuma;1996年)。以下就水稻淀粉生物合成和累积过程中的几个主要酶作一简单介绍。 2.1 ADP葡萄糖焦磷酸化酶(ADP-glucose pyrophosphorylase,AGPP) AGPP是淀粉合成途径中最广泛鉴定的酶,它催化G1P(1磷酸葡萄糖)和ATP形成焦磷酸和 ADPG。ADPG是淀粉生物合成的最初葡萄糖供体,它是一个四聚体,由两个大亚基和两个小亚基组成。AGPP是一个变构调节酶,它正向受3-磷酸甘油的激活,负向受无机磷酸的抑制(Preiss and Sivak,1998年)。在水稻胚乳中,酶活性与淀粉积累是一致的(Anderson等,1989年),与籽粒灌浆速率有很好的相关性(潘晓华等,1999年;杨建昌等,2001年),是淀粉合成的限速酶(Greene,1998年),但不影响淀粉的结构(朱昌兰等,2002年)。 2.2 淀粉合成酶(starch synthase) 淀粉合成酶是一个葡萄糖转移酶,它以寡聚糖为前体,ADPG为底物,通过α-1,4糖苷键不断增加寡聚糖的葡萄糖单位,最终形成α-1,4糖苷键连接的聚糖,聚糖又将作为淀粉分支酶的底物合成支链淀粉。淀粉合成酶依据它在淀粉体中存在的状态,可分为颗粒结合型淀粉合成酶(granule-bound starch synthase,GBSS)和可溶性淀粉合成酶(soluble-starch synthase,SSS)(Forster,1996年)。 水稻胚乳中GBSS是直链淀粉合成过程中起主要作用的酶,由Wx位点控制,该Wx位点的基因产物(Wx蛋白)是60 kDa左右的蛋白,紧密地结合在淀粉粒上。与正常的野生型水稻相比,缺失GBSSⅠ的水稻胚乳中不含直链淀粉(Takeda and Hizukuri,1987年)。王宗阳和武志亮(1991年)及蔡秀铃和王宗阳(1997年)研究表明,不同水稻品种胚乳直链淀粉的含量受Wx基因转录后加工,尤其受第一内含子从前体mRNA切除效应的调控。Sano等(1986年)发现在非糯品种中又可根据Wx蛋白的特性,进一步分为Wxa和Wxb两种等位基因,Wxa和Wxb在不同水稻亚种间已出现明显分化,其中籼稻(包括野生稻)以Wxa为主,直链淀粉含量较高;粳稻全为Wxb,直链淀粉含量较低。包劲松等(2000年)研究水稻Wx基因(CT)n微卫星标记与稻米淀粉品质的关系发现,表观直链淀粉含量、糊化温度、胶稠度和淀粉黏滞性谱等淀粉品质性状都与Wx基因有关,并提出利用微卫星标记进行标记辅助选择可以改良稻米的淀粉品质。此外,GBSS不仅仅涉及到直链淀粉的合成,而且还涉及分离的淀粉颗粒中支链淀粉分支的延长(Denyer等,1996年),但GBSS在正常淀粉粒的支链淀粉链的延长中的确切作用还不清楚。 可溶性淀粉合成酶是另一大类的淀粉合成酶,它包括了除GBSS Ⅰ以外的所有淀粉合成酶,其同工型很多,根据其氨基酸序例可将这些同工型分为3类:SSⅠ、SSⅡ和SSⅢ(Knight等,1998年;Ham等,1998年;Gao等,1998年)。有关这些同工型在淀粉合成中的作用都是渊源于缺失某一同工型的突变体和转基因植物,而这些表现型的植株似乎表明:每种同工型在支链淀粉合成中扮演着特定的作用,如SSⅡ形式缺失,导致支链淀粉中间长度的链减少,短链增加,这表明SSⅡ在中间长度链的合成中有专一作用,其他同工型酶不能代替这种缺失作用(Craig等,1998年)。但每种同工型的可溶性淀粉合成酶在淀粉合成过程的确切作用仍不清楚。 2.3 淀粉分支酶又称Q酶(starch branching enzyme,SBE) SBE在支链淀粉结构形成中有着重要的作用,催化α-1,6糖苷键的形成,与可溶性淀粉合成酶共同作用,形成支链淀粉(Smith等,1995年;Burton等,1995年)。SBE在植物器官中含有多个同工酶(Martin and Smith,1995年)。Martin和Smith(1995年)以及Smith等(1995年)认为同工型淀粉分支酶在结构和功能上的差异决定支链淀粉簇状结构内部分支模式和链长的分配。根据淀粉分支酶作用的底物和形成的分支链长可将同工型分为A型和B型,A型偏向于短的葡萄糖苷链,而B型偏向于长的葡萄糖苷链(Guan and Preiss,1993年)。杨建昌等(2001年)研究表明,水稻籽粒中3个酶(ADPG焦磷酸化酶、淀粉合成酶和淀粉分支酶)的活性变化与籽粒灌浆动态相关联,尤以Q酶的相关值最大,对籽粒灌浆起着关键的调控作用。水稻SBEⅢ缺失突变体表现直链淀粉扩展,它的特征是表观直链淀粉含量格外的高(Kim等,1998年)。李太贵等(1997年)认为高温下早籼以腹白为主的垩白形成主要是包括Q酶在内的酶的绝对或相对的缺乏引起的,糖源不足可能不是主要原因。Nakamura等(1992年)对Q酶进行了较为系统的研究,他认为各种淀粉分支酶活性平衡点的变化,改变了多聚葡萄糖的结构和颗粒形状,这实际上确定了稻米的品质。 2.4 淀粉脱支酶(debranching enzyme,DBE) DBE是水解α-1,6糖苷键的酶。近年来,淀粉脱支酶在支链淀粉结构形成中的作用越来越受到关注。Kubo等(1999年)通过对水稻胚乳sugary-1等位基因研究,表明淀粉脱支酶即异淀粉酶(isoamylose,ISA)和普鲁兰酶(pullanase)都参与了水稻胚乳支链淀粉的生物合成。其他研究也证实了这一观点(Nakamura等,1996年;Afroza,1998年;David,2000年)。水稻胚乳中R-enzyme(RE)的分子大小在100~105 kDa(Nakamura等,1996年;Toguri,1991年)。到目前为止,有关淀粉脱支酶在支链淀粉合成中的作用有两种假设,第一种,是以Erlander(1970年)为代表的,认为植物糖原是支链淀粉合成的中间产物,经过脱支酶的作用形成支链淀粉;另一种是以Nakamura和Yuki(1992年)和Nakamura等(1996年)为代表,认为分支酶和脱支酶这两种酶的酶活性平衡对α-1,6-分支的频率或支链淀粉的α-1,4侧链的链长的分配有着重要的决定作用。这两种假定都各有欠缺,还有待进一步研究论证。 因此,支链淀粉的合成和淀粉颗粒的形成是一个复杂的且可调节的过程,它是由SS、SBE以及淀粉其他代谢酶共同作用的结果。 3 淀粉粒的形态与结构及其糊化特性 3.1 淀粉粒的形态 淀粉粒的大小及形状也反映籽粒品质的优劣。品质较好的水稻品种,籽粒中淀粉粒较小,呈明显的多面体晶形,棱角明显且清晰可见,排列整齐且紧密,粒间隙极小;而品质较差的籽粒中及垩白部位的淀粉粒,颗粒较大且多面体的棱角不明显,个别近圆形,粒问疏松(符文英等,1997年)。季清娥等(1998年)也证实了这一点,但又发现劣质品种具垩白,其淀粉粒有两种形状,非垩白处淀粉粒为大小一致的多面体,排列紧密,垩白处淀粉粒棱角不明显或呈圆球形,大小参差不齐,受压力时可见单个淀粉粒和小球形颗粒。吴殿星等(2001年)研究发现云雾性状是低表观直链淀粉含量品种(系)特有的胚乳外观,其出现的临界表观直链淀粉含量上限可能在14.5%左右,而下限为9%~10%。 此外,胚乳细胞的分裂及生长状况对籽粒品质有一定的影响。范燕萍等(1989年)研究表明胚乳细胞的数目、大小及其形状与垩白性状有密切的关系,籽粒中分裂形成的小细胞越多,米粒垩白越大,垩白率越高。符文英等(1997年)研究表明,横断面胚乳细胞的长宽比与米质关联度存在极显著负相关,细胞细长的米质都较差,细胞较粗短的米质都较优;淀粉粒的粒径变异系数与综合米质指标存在极显著负相关,淀粉粒的大小不均匀是导致米质性状下降的一个主要原因,并提出淀粉粒粒径变异系数可作为衡量米质优劣的间接指标。王忠等(2003年)认为,离背部维管束远的腹部胚乳细胞因养分输导途径长,易充实不良,形成腹白米,大粒品种的中部的胚乳细胞由于其离糊粉层也相对较远,也易形成心白。有关垩白性状受环境条件的影响曾有多次报道(赵式英,1982年;李太贵等,1997年;蔡一霞等,2002年),但是这种影响是否是通过环境对胚乳细胞的数目、大小及其形状的影响来实现的,目前还未见这方面的报道。 3.2 淀粉的结构 稻米淀粉以复粒淀粉形式贮藏于胚乳淀粉体中。单个淀粉粒是由两部分组成,其中一部分为直链淀粉(amylose),是以α-1,4糖苷链连接而成的数千个单位长的极少分支的葡萄糖链状分子,具有热水溶性,另一部分是支链淀粉(amylopectin),不溶于热水,是由短的α-1,4糖苷链相连葡萄糖通过的α-1,6糖苷链连接而成的高度分支的葡萄糖聚合物,大约每10~20个葡萄糖单位就一个分支,分支的链长为12~60个葡萄糖单位。在天然的大米淀粉颗粒中,直链淀粉不能形成结晶,而是以单螺旋结构掺入支链淀粉分子形成的疏密相间的结晶区和无定形区间。淀粉粒的晶体结构实质上是支链淀粉外部的线性链以双螺旋结构存在于晶体的片层中(图2)。 以往大量的研究表明,直链淀粉含量直接影响稻米的食用蒸煮品质(Juliano and Villareal,1993年)。这种观念一直延续到20世纪80年代中期,直链淀粉和支链淀粉比例一直被大多数人认为是决定稻米蒸煮食用品质的主要因素,因此国内外将直链淀粉含量作为衡量稻米品质的一个主要指标。但近来的研究发现,直链淀粉含量相近的品种之间(尤其是中等和高等直链淀粉含量品种)米饭质地表现出明显的差异,并认为直链淀粉含量并非对米饭质地具有完全的决定作用。Bhahacharya等(1982年)认为表观直链淀粉中不溶的部分是米饭质地主要的决定因素。Chinnaswany和Bhattacharya(1986年)用凝胶渗透色谱技术(以sepharose 2B为填料)分离水稻淀粉,发现淀粉中分子量大的有分支的部分(FRⅠ)与水稻不溶性的直链淀粉有着密切的关系,而分子量小的部分(FRⅡ)与直链淀粉的可溶部分有着密切关系。随后Reddy等(1993年)用GPC(gel-permeation chromatography)分离FRⅠ也证实了这一点,并认为热水不溶的直链淀粉实质是支链淀粉分支链中的长链B部分,正是由于这些长链B分子间的相互作用,使得淀粉颗粒强硬和有弹力,从而导致米饭质地硬,而短链部分多,米饭就越软。Ong和Blanshard(1995年)研究表明支链淀粉中长链越多且短链少的水稻品种,其米饭质地就越硬,反之,支链淀粉中长链越少且短链越多的水稻品种,其米饭质地就越软。而Ramesh等(1999年)认为淀粉中所有的线性长链包括直链淀粉本身控制着米饭的质地。 直链淀粉是由α-1,4糖苷键连接而成的线性葡萄糖聚合物,是在淀粉合成酶异构体GBSSⅠ(颗粒结合淀粉合成酶)的作用下形成的。目前有3个方面的研究现象表明,体内直链淀粉的合成与淀粉粒矩阵的形成交织在一起(1)随着GBSSⅠ活性的减少,颗粒内直链淀粉的分配不均;(2)在分离的淀粉颗粒中,经GBSSⅠ作用形成的是支链淀粉,而不是直链淀粉;(3)某些物种的淀粉合成速率与直链淀粉含量成显著正相关。在研究马铃薯淀粉合成过程中,通过增加GBSSⅠ的活性,其直链淀粉含量的增加不超过20%,这表明直链淀粉最大值取决于支链淀粉矩阵内所提供的空间。Smith等(1997年)认为贮藏器官中有以下4个因素决定直链淀粉合成速率和直链淀粉含量:①GBSSⅠ蛋白的数量;②ADPG供应能力;③麦芽寡糖的供应能力;④支链淀粉合成所提供的矩阵空间的可用性。这4种因素相对的重要性因贮藏器官的不同而不同,但是在野生型贮藏器官中矩阵空间的可用性是直链淀粉含量的最终决定因素。 3.3 淀粉糊化特性 淀粉的生理生化特性包括了淀粉的颗粒性状、糊化特性和黏度特性等。在水稻品质的研究过程中,目前研究稻米淀粉生理生化特性最多的是稻米的糊化特性。水稻淀粉(或米粉)溶于水时,只形成悬浮液,当悬浮液被加热到一定温度时,颗粒开始剧烈膨胀,颗粒外围的支链淀粉被胀裂,内部的直链分子游离出来,悬浮液变成黏稠状,这种现象称为淀粉的糊化。淀粉粒开始剧烈膨胀时的温度称为糊化温度。水稻淀粉的糊化温度因品种的不同或淀粉粒大小不同,其糊化温度也有区别。由于破裂的支链淀粉在糊浆中形成凝胶与流释出来的直链淀粉在糊浆中形成溶胶的黏度之间的差异,淀粉在糊化过程中,其淀粉糊的黏度发生一系列的变化。黏度速测仪(rapid viscosity analyzer,RVA)和差示扫描量热法(differential scanning carolina,DSC)常用于分析淀粉黏度和热变化的仪器和方法(Reddy等,1994年;Juliano,1996年;Okadome等,1998年)。大量研究表明,稻米淀粉糊化特性与稻米品质有着密切的关系,并在稻米育种中发挥着重要的作用(吴殿星等,1998年;200l年)。吴殿星等采用黏度速测仪分析不同早籼品种,发现消减值和崩解值等黏滞谱特征值,能特异性区分不同品种表观直链淀粉含量(AAC)的高低和鉴定中等AAC品种食用品质的优劣,糯稻和低AAC品种的消减值为负值,中等AAC品种消减值为正值;崩解值大的,胶稠度较好,消减值与米饭的质地和口感相关联,消减值为正值且较小时,米饭表观软而不黏,口感好,正值过大时,米饭硬而糙,但消减值为负值时,米饭往往表现过黏。金正勋等(2001年)也证实了这一点。包劲松等(1999年)发现稻米的RVA谱特性主要受Wx基因控制,认为将粳稻Wx基因引入籼稻背景中可改良稻米淀粉RVA谱。 4 蛋白质和脂类与稻米品质的关系 大米中的蛋白质含量为6%~8%,在谷类作物中属于低值,但在生物体中的利用率比其他谷类要优越,其质量最好。因此,蛋白质是稻米营养品质的重要指标。但有研究表明,蛋白质与稻米食味呈负相关关系,蛋白质含量高,米饭的食味就越差(张国民等,2001年)。事实上,稻米蛋白中的白蛋白、球蛋白和谷蛋白等都是由一些优良氨基酸组成,其营养丰富而不影响食味的蛋白质;只有阻碍淀粉网眼状结构发展的醇溶谷蛋白,才是导致食味降低而又几乎不为肠胃所吸收的蛋白质;其次稻米中游离氨基酸是提高食味的成分,但其前体物质酰胺以及铵离子则是降低食味的因素(孙平,1998年)。 稻米中的脂类的含量很少,糙米中仅含2.4%~3.9%。虽然它是稻米营养成分之一,但它的组成对稻米食味品质有着很大的影响。Marshall等(1990年)发现,脂类和蛋白质对稻米淀粉糊化有不可忽视的影响。Tester和Morrison(1990年)在研究支链淀粉、直链淀粉和脂类对谷物淀粉膨润和糊化的影响时指出,淀粉膨润是支链淀粉的特性,而直链淀粉只起稀释剂的作用,但天然淀粉中的直链淀粉和脂类形成复合物时就起到抑制淀粉膨润的作用。刘宜柏等(1982年)对早籼稻品质的米质进行相关性分析后表明,稻米脂类含量较其他组分对稻米食味品质有更大的影响,脂肪含量高,直链淀粉含量中等偏低,胶稠度软或中等偏软,米粒延伸性好的稻米食味品质好。脂肪含量越高的稻米,米饭光泽越好。 5 结语 综观稻米品质的研究,无论在育种上或在栽培上均取得了较大的进展。从前人的研究中不难发现,稻米品质的形成是受遗传和环境二者共同作用的,而环境条件对稻米品质的影响也是通过影响稻株和颖果的生理过程而发挥作用的(王忠等,2003年)。稻米品质的形成涉及到籽粒灌浆过程中"源、库和流"3个方面的协调作用,实质上是米质形成过程中碳、氮及脂肪代谢过程。因此,只要是对这些过程有影响的因子均可能影响到米质的形成。从已有的内容来看,稻米品质研究的一个共同点就是集中在成熟期稻米品质表现的有关性状的分析上,而对品质形成的机理及米质形成的过程中生理生化特性与米质优劣有何关系的研究相对较少,有关淀粉、蛋白质及脂类研究多局限在量上,与食味相关的分子结构研究更为鲜见。因此,研究结果对指导米质改良的效果不甚明显,且在稻米种植的调优措施上也显得比较薄弱,这使得我国的稻米品质研究与国际上近十年研究成果差距较大。因此,可通过加强稻米品质生理基础的研究,从生理角度来了解稻米品质形成的内在机理,对优质育种和优质米栽培有着重要的意义。 注: (浏览次数:4115)
|
| 上篇文章 | 下篇文章 | 相关文章 | 推荐给朋友 | 打印 | 关闭窗口 |
| 免责声明: |
|
| Copyright © 2003 CNRRI. All rights reserved. 中国水稻研究所 版权所有 地址:杭州市体育场路359号(邮政编码:310006) E-mail:[email protected] |
{kind=link}
{kind=link}